AntibodySystem Supports the Discovery of Hyperoside Targeting DHX9 to Resolve R-Loops in Unexplained Recurrent Pregnancy Loss
Unexplained recurrent spontaneous abortion (URSA) frustrates clinicians and patients alike: after excluding known etiologies, nearly half of all cases remain idiopathic. Emerging evidence points to premature senescence of endometrial stromal cells as a key driver of decidualization failure, but the upstream trigger has been unclear. Now, a study in iMeta(IF: 44.4) titled "Hyperoside alleviates endometrial stromal cell senescence in unexplained recurrent spontaneous abortion via DHX9‑mediated R‑loop resolution" provides an answer. The team found that aberrant R‑loop accumulation in stromal cells ignites a cGAS‑STING‑driven senescence cascade, and that hyperoside-a flavonol glycoside from edible plants like St. John's wort and hawthorn-breaks this cascade by directly targeting DHX9, rescuing decidualization and reducing embryo loss.


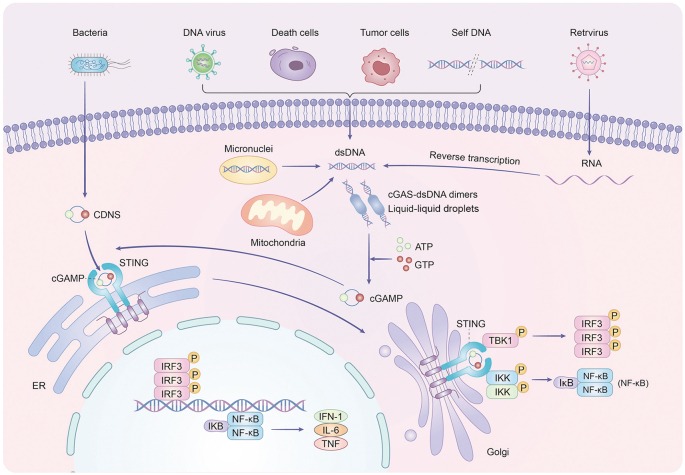
The hidden threat: R‑loops as genomic arsonists
R‑loops are three‑stranded nucleic acid structures composed of an RNA:DNA hybrid plus a displaced single‑stranded DNA (ssDNA). Under physiological conditions, they assist transcription and immunoglobulin switching. But when they accumulate uncontrollably, they become dangerous: they collide with replication forks, induce DNA double‑strand breaks, and-critically-release cytosolic ssDNA fragments that are potent agonists of the cGAS-STING innate immune pathway.
Once activated, cGAS-STING ignites two pro‑senescent branches: IRF3‑driven type I interferon production and NF‑κB‑mediated SASP (senescence‑associated secretory phenotype) secretion. Together with DNA damage markers (γH2AX) and cell‑cycle inhibitors (p53, Rb), this signaling network pushes stromal cells into an irreversible senescent state, crippling their ability to decidualise and support pregnancy. Although R‑loop pathology has been implicated in cancer and neurodegeneration, its role in URSA had never been explored-until now.

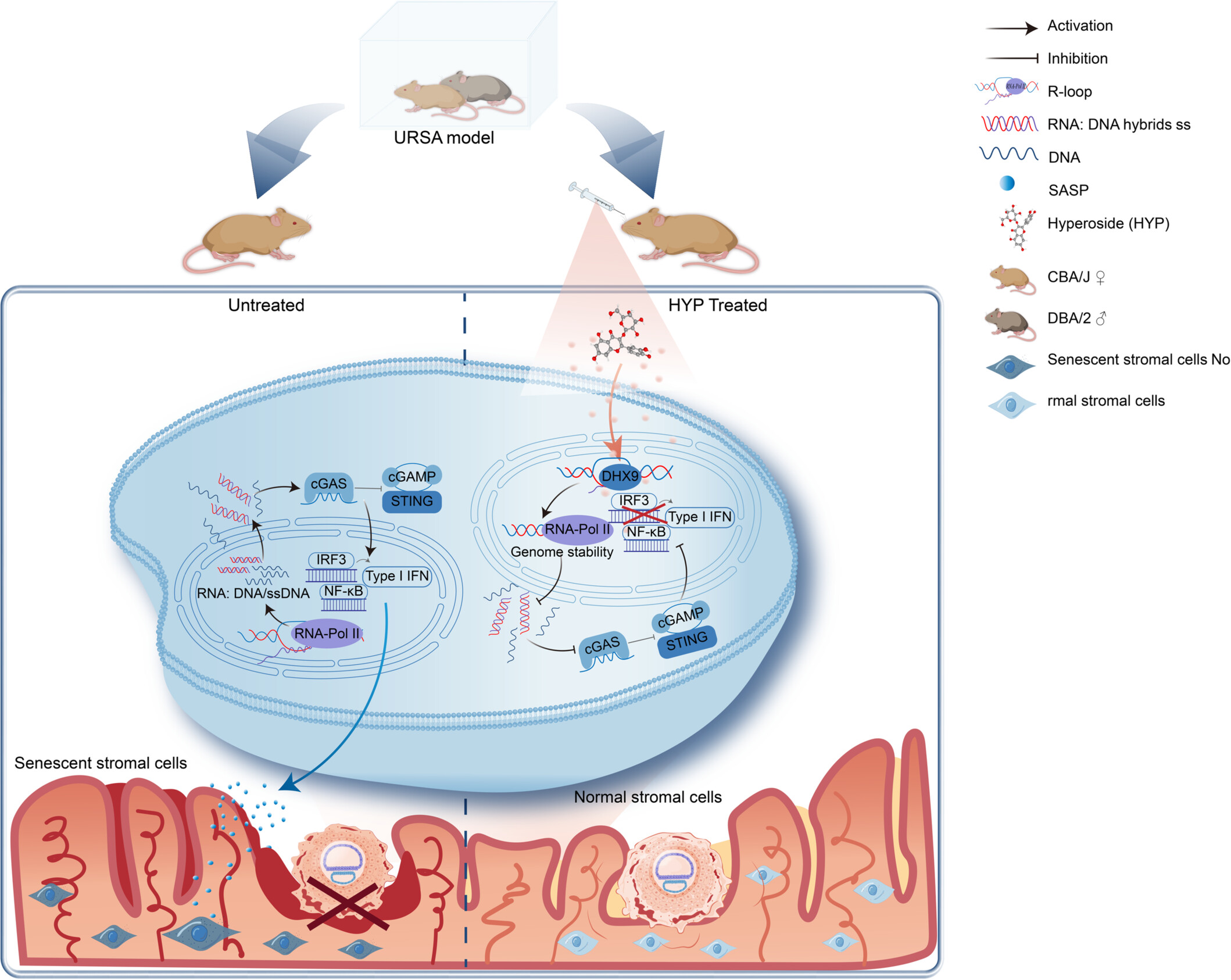
Hyperoside clears R‑loops and rescues decidualization
The team first confirmed that URSA decidual tissues exhibited markedly elevated R‑loop signals, detected using the Anti-DNA-RNA Hybrid Antibody (S9.6) (RGK60001, AntibodySystem), along with increased cGAS and STING fluorescence, relative to normal pregnant controls. These clinical observations prompted mechanistic investigation.
Using the CBA/J × DBA/2 mouse model of URSA, oral hyperoside (9, 18, or 36 mg/kg) produced dose‑dependent protection. High‑dose hyperoside significantly reduced embryo resorption rates, restored the decidual markers PRL and IGFBP1, and normalised stromal cell alignment, collagen deposition, and F‑actin cytoskeletal organisation. Notably, although the standard drug dydrogesterone improved tissue morphology and resorption, it failed to clear R‑loops or lower senescence markers-highlighting hyperoside's unique action at the very beginning of the pathogenic chain.

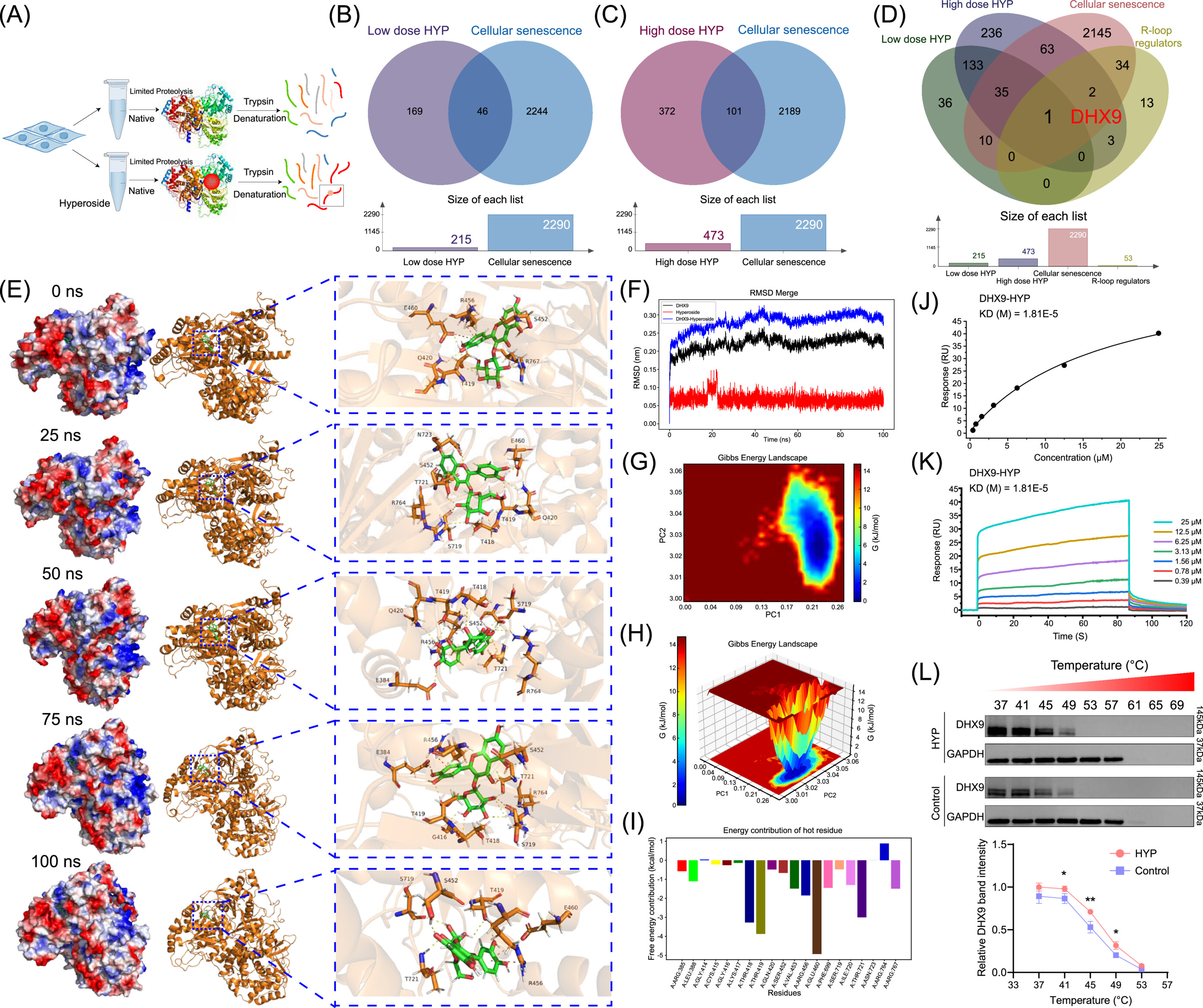
DHX9 at Thr419: the molecular anchor
To pinpoint how hyperoside eliminates R‑loops, the team applied limited proteolysis-mass spectrometry (Lip‑MS) on primary URSA stromal cells, followed by molecular docking, dynamics simulations, surface plasmon resonance (SPR), and cellular thermal shift assays (CETSA). They identified DHX9 (RNA helicase A) as the functional target, with the Thr419 residue (Thr421 in mice) forming a critical hydrogen bond with hyperoside.
SPR measurements revealed a dissociation constant (KD) of 1.81×10⁻⁵ M for wild‑type DHX9, whereas the T419A mutant showed virtually no binding (KD > 2.47 M). CETSA further confirmed that hyperoside binding enhanced DHX9 thermal stability. The authors acknowledge that the exact allosteric mechanism by which hyperoside boosts DHX9 activity remains to be fully defined a point they candidly discuss as a current limitation.

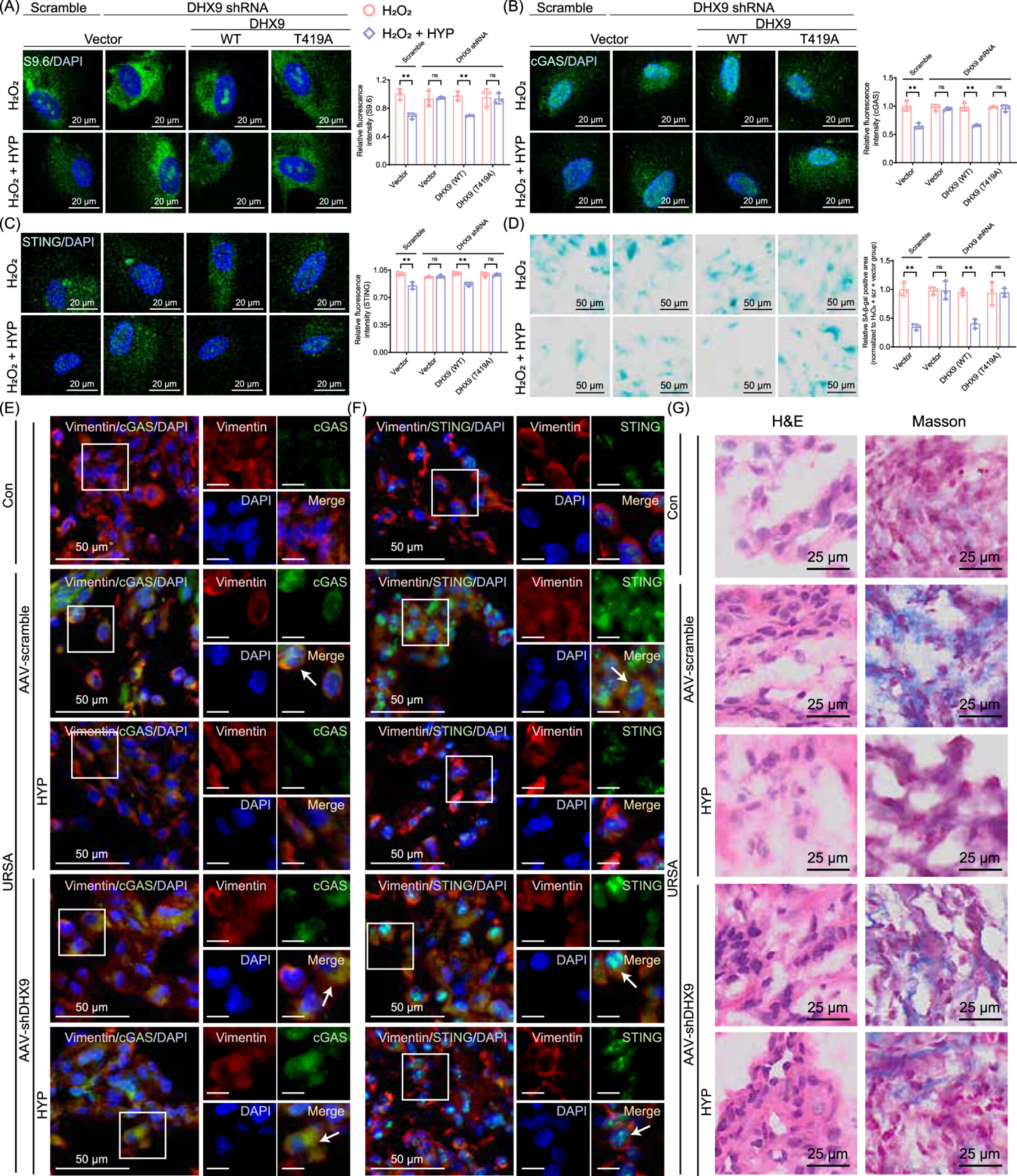
DHX9 is indispensable for hyperoside's therapeutic effect
The final piece of functional validation was decisive. Uterine‑specific DHX9 knockdown via AAV in URSA mice almost completely abolished hyperoside's benefits: R‑loops persisted, cGAS-STING remained activated, senescence markers stayed high, and decidualisation failed to recover. Conversely, adenoviral reconstitution of wild‑type DHX9 restored hyperoside's full therapeutic efficacy, while reconstitution with the T421A mutant (mouse equivalent of T419A) did not. These findings firmly establish that hyperoside's action is strictly dependent on its precise engagement with DHX9.

A paradigm shift in URSA therapy
Collectively, this work delineates a complete pathogenic axis-R‑loop accumulation → cytosolic ssDNA → cGAS-STING activation → stromal senescence → decidualisation failure-and shows that hyperoside disrupts this axis at the very first step by targeting DHX9. This reframes URSA not merely as a hormonal imbalance but as a disorder of transcription-replication conflict resolution, opening a new frontier for therapeutic intervention.
Hyperoside, as a natural compound already present in common foods, offers a dual advantage: it stabilises the genome and curbs inflammation, addressing two facets of URSA that progesterone cannot touch. While clinical translation will require thorough pharmacokinetic and safety evaluations, the study provides a clear molecular target (DHX9 Thr419) and a natural lead scaffold for future drug development. The authors also suggest that screening DHX9 polymorphisms in URSA cohorts might one day guide personalised dietary recommendations based on hyperoside‑rich foods.
Reagents and resources
Beyond S9.6, AntibodySystem offers a comprehensive portfolio of recombinant antibodies targeting diverse nucleic acid structures, including ssDNA, dsDNA, hairpin DNA, Z‑DNA, i‑Motif, G‑tetramer, dsRNA, and various RNA species.
| Catalog No. | Product Name |
|---|---|
| RGK60001 | Anti-DNA-RNA Hybrid Antibody(S9.6) |
| RGK60301 | Anti-DNA/RNA G-quadruplex Structures Antibody (BG4) |
| RGK60307 | Anti-DNA/RNA G-quadruplex Structures Antibody (BG4) |
| RGK60305 | Anti-G-quadruplex DNA Antibody (4E11#) |
| RGK60306 | Anti-G-quadruplex DNA Antibody (1H6) |
| RGK60308 | Anti-G-quadruplex DNA Nanobody (SAb2479) |
| RGK24020 | Anti-dsDNA Antibody (3E10#) |
| RGK24021 | Anti-dsDNA hIgG Antibody (3E10#) |
| RGK23954 | Anti-B-DNA Antibody (Fs0492) |
| KAK24001 | Anti-dsDNA Mouse IgG ELISA Kit |
| KGK24001 | dsDNA ELISA Kit |
| RGK24901 | Anti-dsRNA Antibody (1D3) |
| RGK24902 | Anti-dsRNA Antibody (10G1) |
| RGK24903 | Anti-dsRNA Antibody (9D5) |
| RGK24904 | Anti-dsRNA Antibody (6G9) |
| RGK24905 | Anti-dsRNA Antibody (10B2) |
| RGK24910 | Anti-dsRNA Antibody (SAb2564) |
| RGK24906 | Anti-dsRNA Antibody (J2) |
| RGK24907 | Anti-dsRNA Antibody (K1) |
| RGK24908 | Anti-dsRNA Antibody (K2) |
| KGK24901 | dsRNA ELISA Kit |
| RGK24118 | Anti-ssDNA Antibody (S1-15) |
| RGK25001 | Anti-ssRNA Antibody (JEL103) |
| RGK25002 | Anti-ssRNA Antibody (BRG) |
| RGK23911 | Anti-DNA Antibody (BKV29) |
| RGK23912 | Anti-DNA Antibody (BKV39) |
| RGK23916 | Anti-DNA Antibody (B8801) |
| RGK24014 | Anti-DNA Antibody (50) |
| RGK24801 | Anti-RNA Antibody (D444) |
| RGK08001 | Anti-RNA Antibody (D44) |
| RGK08003 | Anti-RNA Antibody (Bl3-6) |
| RGK24301 | Anti-Z-DNA Antibody (Z44) |
| RGK24602 | Anti-Triplex DNA Antibody (Jel318) |
| RGK24601 | Anti-Triplex DNA Antibody (Jel466) |
| RGK24501 | Anti-Hairpin ssDNA Antibody (DNA-1) |
| RGK23940 | Anti-ssDNA/dsDNA/Z-DNA Antibody (B3) |
| RGK24202 | Anti-dsDNA/ssDNA Antibody (4B2) |
| RGK24201 | Anti-dsDNA/ssDNA Antibody (A52) |
| RGK23951 | Anti-dsDNA/ssDNA Antibody (3D8) |
| RGK24302 | Anti-Z-DNA/Z-RNA Antibody (Z22) |
| RGK24909 | Anti-dsRNA/ssRNA Antibody (P6) |



